'Urmakerens blindhet'
(Fritt oversatt fra 'Not By Chance; kap. 6, av Dr. Lee Spetner -{oversetters merknader i klamme-parentes})
 I 1986 publiserte zoologen R. Dawkins boken 'The Blind Watchmaker'. Dawkins har gitt uttrykk for at livet har utviklet seg helt av seg selv, og ønsket med misjonerende iver å gi denne overbevisningen videre. Han argumenterte at Neo-Darwinistisk Teori (NDT) godt forklarte hvordan livet oppsto og utviklet seg, og at det ikke var noen grunn til å kalle på noen skaper i den sammenheng. I boka tok Dawkins utgangspunkt i W. Paleys Argument For Design (AFD) som trekker en analogi fra opphavet på ei klokke til levende organismer. AFD starter med å slutte til eksistensen av en urmaker, ut fra eksistensen av et ur. Paley-argumentet brukes til å uttrykke en konklusjon, i det det er praktisk umulig for et livløst objekt å bli til av tilfeldighet. Det samme gjelder for livet i følge AFD.
I 1986 publiserte zoologen R. Dawkins boken 'The Blind Watchmaker'. Dawkins har gitt uttrykk for at livet har utviklet seg helt av seg selv, og ønsket med misjonerende iver å gi denne overbevisningen videre. Han argumenterte at Neo-Darwinistisk Teori (NDT) godt forklarte hvordan livet oppsto og utviklet seg, og at det ikke var noen grunn til å kalle på noen skaper i den sammenheng. I boka tok Dawkins utgangspunkt i W. Paleys Argument For Design (AFD) som trekker en analogi fra opphavet på ei klokke til levende organismer. AFD starter med å slutte til eksistensen av en urmaker, ut fra eksistensen av et ur. Paley-argumentet brukes til å uttrykke en konklusjon, i det det er praktisk umulig for et livløst objekt å bli til av tilfeldighet. Det samme gjelder for livet i følge AFD.
Bilde 1. Boka 'The Blind Watchmaker' 1.utg
Dawkins argument var at levende organismer som kan reprodusere seg, kan utvikles gradvis via seleksjon fra noe enklere. Sjansen for at dette skal skje av seg selv, er mye større enn at alt skal komme sammen samtidig. Dawkins konkluderte at NDT, postulatet om naturlig variasjon fulgt av naturlig seleksjon, fornekter AFD. I følge Dawkins var livet et resultat av sjanse, heller enn design. Hans analogi til urmakeren, er ikke skaperen som Paley hevdet, der er blind tilfeldighet eller som han uttrykte det' urmakeren er blind'. Dawkins er hva forfatteren har kalt en gradvis evolusjonist. Han mener at makroevolusjon gradvis finner sted, gjennom en lang serie av små steg (kumulativ seleksjon). Hvert ledd i kjeden er en liten tilpasset endring, fulgt av naturlig seleksjon. Vi har tidligere vist at sjansen for at kumulativ seleksjon skal være byggesteinen i livets utvikling, er for liten til at det skjer i praksis.
Selv om Dawkins skrev om tilfeldighet, beregnet han ikke sjansen for noe som helst. {Han har tvert om i andre sammenhenger senere uttalt: " Det er knasende, knirkende og kjedelig innlysende at hvis darwinismen virkelig var en teori som handlet om tilfeldigheter, dugde den ikke. Man trenger ikke være matematiker eller fysiker for å regne ut at det ville vare herfra til en mindre evighet før et øye eller et hemoglobin-molekyl ville kunne sette seg selv sammen ved hjelp av rent og skjært hulter-til-bulter hell"(1). Sir Fred Hoyle og astrofysiker Chandra Wixkramasingh deler Dawkins mening i denne henseende. "Uansett hvor stort miljø en trekker inn i ligningen, kan ikke livet være begynt ved ren og skjære tilfeldigheter. Sannsynligheten for livets spontane dannelse fra død materie, er 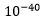 (1-tall på plass 40 bak desimalpunktum). Et slikt utgangspunkt gjør at det aldri har vært noen 'ursuppe' på jorden eller andre steder. Hvis livet begynnelsesstadier ikke var tilfeldige, må de ha vært produkt av en målrettet intelligens. (2).
(1-tall på plass 40 bak desimalpunktum). Et slikt utgangspunkt gjør at det aldri har vært noen 'ursuppe' på jorden eller andre steder. Hvis livet begynnelsesstadier ikke var tilfeldige, må de ha vært produkt av en målrettet intelligens. (2).
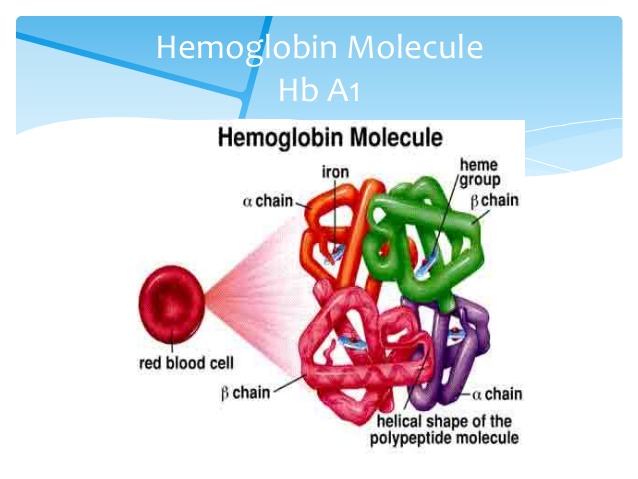
Dawkins innrømte at sjansen for at en sjanse på 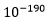 (10 opphøyd i -190) var for liten til at hemoglobin kunne bli til i ett steg. Men han gjorde ikke annet enn å påpeke at sjansen for at det kunne oppstå ved kumulativ seleksjon var større. Han foreslo ikke hva som kunne være en akseptabel sannsynlighet. Da vi i kap 4 så på sjansen for forekomst av de små stegene som evolusjon skulle bygge på, fant vi at den var liten om ikke to presise forutsetninger er tilfredsstilt: i) De må ha en selektiv fordel og ii) I gjennomsnitt må de tilføre en liten informasjonsmengde til genomet.
(10 opphøyd i -190) var for liten til at hemoglobin kunne bli til i ett steg. Men han gjorde ikke annet enn å påpeke at sjansen for at det kunne oppstå ved kumulativ seleksjon var større. Han foreslo ikke hva som kunne være en akseptabel sannsynlighet. Da vi i kap 4 så på sjansen for forekomst av de små stegene som evolusjon skulle bygge på, fant vi at den var liten om ikke to presise forutsetninger er tilfredsstilt: i) De må ha en selektiv fordel og ii) I gjennomsnitt må de tilføre en liten informasjonsmengde til genomet.
Bilde 2. Hemoglobin i røde blodceller
Vi har sett atskillige eks. på at forutsetningene ikke er tilfredsstilt, og det foreligger ikke eks. der forutsetningene er tilfredsstilt. Vi må konkludere med at Dawkins tok feil i det han skrev om kraften ved kumulativ seleksjon. Dawkins gjorde en avgjørende feil som ødela for konklusjonen hans. Han antok at stegene som utgjør kumulativ seleksjon kunne være så små en kunne ønske. Med dette henførte han til at de kunne være så sannsynlig å hende som en kunne ønske. Denne forutsetningen er feil. Kravet om selektiv fordel hindrer at stegene kan være så små en ønsker. Mens Dawkins viet et kap. til 'kraften av kumulativ seleksjon', så viet han ingen plass til hvorvidt slik kumulativ seleksjon var mulig. Til det må en beregne sannsynligheter.
Mutasjoner kan ikke være så små en ønsker. Forfatterens analogi var å prøve å forbedre en novelle ved å bytte ut enkeltbokstaver. Men skulle det være noen sammenheng i endringene, så ville det kreve andre endringer i tillegg. Og fordi hvert steg må tilfredsstille noen minimumskrav, så kan en ikke bare anta at endringene kan være vilkårlig små. Deres minimum størrelse, er en punktmutasjon i DNA-nukleotiden -som må følges av tilsvarende endringer andre steder om det skal ha noen selektiv verdi. Det er mye vi fremdeles ikke vet om levende organismer som lever i populasjoner, om deres samvirke og reaksjon på ytre press mot populasjonen. Men vi kan ikke bruke vår uvitenhet som lisens for å skape kunstige fakta for å støtte en favoritt-teori.
 Dawkins tok mål av seg til å forklare at livets kompleksitet er et resultat av naturlige lover. Han befattet seg da også med livets opprinnelse. Han bemerket at kumulativ seleksjon bare kunne virke på systemer som kan reprodusere seg selv. Hvordan kunne han så få til opprinnelsen til det første levende system? Det kunne i hvert fall ikke ha skjedd ved kumulativ seleksjon. {Vi vet fra andre sammenhenger senere, at Dawkins faktisk ikke ser helt bort fra en intelligens ved livets opprinnelse, men det må bare ikke være en skaper her på jorda..-oversetters merknad. Vi velger dermed å se bort fra den ikke-overbevisende argumentasjonen i 'The Blind watchmaker' på dette punktet, slik Dawkins selv synes å ha gjort.} Det tyder på at han har vært inne på 'alien-tanken' allerede da, siden han foreslår at vi skulle tenke på sannsynligheter i lys av en 'hypotetisk alien' som lever i millioner av år. Han tar ikke inn over seg at en begivenhet som har en gitt sannsynlighet, har denne uansett hvor lang tid en forholder seg til. Han bruker eks. med å få utdelt en perfekt bridgehånd, der hver spiller får utdelt 13 kort av samme slag. Han påstår at en kan forvente at en 'alien som lever en million år' kan forvente å få utdelt en perfekt bridge-hånd fra tid til annen. Men en kan greit kalkulere sannsynligheten for at Dawkins alien får tildelt en perfekt bridge hånd. Sjansen for det er
Dawkins tok mål av seg til å forklare at livets kompleksitet er et resultat av naturlige lover. Han befattet seg da også med livets opprinnelse. Han bemerket at kumulativ seleksjon bare kunne virke på systemer som kan reprodusere seg selv. Hvordan kunne han så få til opprinnelsen til det første levende system? Det kunne i hvert fall ikke ha skjedd ved kumulativ seleksjon. {Vi vet fra andre sammenhenger senere, at Dawkins faktisk ikke ser helt bort fra en intelligens ved livets opprinnelse, men det må bare ikke være en skaper her på jorda..-oversetters merknad. Vi velger dermed å se bort fra den ikke-overbevisende argumentasjonen i 'The Blind watchmaker' på dette punktet, slik Dawkins selv synes å ha gjort.} Det tyder på at han har vært inne på 'alien-tanken' allerede da, siden han foreslår at vi skulle tenke på sannsynligheter i lys av en 'hypotetisk alien' som lever i millioner av år. Han tar ikke inn over seg at en begivenhet som har en gitt sannsynlighet, har denne uansett hvor lang tid en forholder seg til. Han bruker eks. med å få utdelt en perfekt bridgehånd, der hver spiller får utdelt 13 kort av samme slag. Han påstår at en kan forvente at en 'alien som lever en million år' kan forvente å få utdelt en perfekt bridge-hånd fra tid til annen. Men en kan greit kalkulere sannsynligheten for at Dawkins alien får tildelt en perfekt bridge hånd. Sjansen for det er 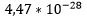 (4.47 *10 opphøyd i -28). Om Dawkins alien får utdelt 100 bridge hender hver dag i sitt liv for 100 millioner år, så ville han få ca.
(4.47 *10 opphøyd i -28). Om Dawkins alien får utdelt 100 bridge hender hver dag i sitt liv for 100 millioner år, så ville han få ca. 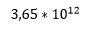 (3,65*10 opphøyd i 12) 'hender'. Sjansen for å få en perfekt hånd minst én gang i livet, er dermed
(3,65*10 opphøyd i 12) 'hender'. Sjansen for å få en perfekt hånd minst én gang i livet, er dermed 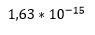 (1,63*10 opphøyd i-15) (ca. 1 til en billiard).
(1,63*10 opphøyd i-15) (ca. 1 til en billiard).
Det ville være noe å skrive hjem om, om en spiller får det.
Bilde 3. Livets opprinnelse -en sammensatt affære
Dawkins feil er en typisk feil som mange evolusjonister gjør. De tror at jordens alder er lang nok til at hva som helst kan inntreffe. En skulle i slike sammenhenger prøve å multiplisere sannsynligheten for at det skjedde én gang med antall forsøk. Denne prosedyren er bare (noenlunde) korrekt om resultatet viser seg å være mindre enn 1. {Ellers så vil det være selvmotsigende, og en har med avhengige begivenheter, som må beregnes med betingede sannsynligheter-oversetters kommentar}. Nødvendige begivenheter for kumulativ seleksjon er altfor usannsynlige å bygge en teori på. Begivenhetene som trengs for livets opprinnelse er enda mer usannsynlige.
Dawkins ga resultatet av noen computer-simuleringer han gjorde i forhold til evolusjon. I boka beskriver han to simuleringer i så måte.  En av disse var simuleringen av en tekst fra Shakespeare. Den andre var utvikling av et sett tegninger. Vi skal se i hvilken grad disse simuleringene kan tas som realistiske. La oss se om vi kan konkludere noe fra dem om NDT. I den 1. simuleringen startet han med en streng av tilfeldige bokstaver og lot den 'utvikle seg' mot en mål-streng. Målstrengen han valgte var setningen i Hamlet: 'ME THINKS IT LOOKS LIKE A WEASEL.' Inklusive mellomrom (uten punktum) har strengen 28 tegn. Han valgte karakterene i startstrengen tilfeldig, og brukte bare STORE BOKSTAVER for å gjøre det enkelt. Han kopierte startstrengen noen ganger for å få strenger til neste generasjon. Hver gang han kopierte den, introduserte han en forutbestemt sannsynlighet for tilfeldig feil. Kopieringer var ment å simulere reproduksjon til levende organisme, og feilene var ment å simulere mutasjoner. Ut fra de varierende resultatene i 2.generasjon valgte computeren ut den som var mest lik målsekvensen.
En av disse var simuleringen av en tekst fra Shakespeare. Den andre var utvikling av et sett tegninger. Vi skal se i hvilken grad disse simuleringene kan tas som realistiske. La oss se om vi kan konkludere noe fra dem om NDT. I den 1. simuleringen startet han med en streng av tilfeldige bokstaver og lot den 'utvikle seg' mot en mål-streng. Målstrengen han valgte var setningen i Hamlet: 'ME THINKS IT LOOKS LIKE A WEASEL.' Inklusive mellomrom (uten punktum) har strengen 28 tegn. Han valgte karakterene i startstrengen tilfeldig, og brukte bare STORE BOKSTAVER for å gjøre det enkelt. Han kopierte startstrengen noen ganger for å få strenger til neste generasjon. Hver gang han kopierte den, introduserte han en forutbestemt sannsynlighet for tilfeldig feil. Kopieringer var ment å simulere reproduksjon til levende organisme, og feilene var ment å simulere mutasjoner. Ut fra de varierende resultatene i 2.generasjon valgte computeren ut den som var mest lik målsekvensen.
Bilde 4. Hvor mye tilfeldig kan skje i universets levetid
Computerens valg av den beste 'kopieringen' var ment å simulere naturlig seleksjon. Fra den utvalgte strengen foredlet computeren 3. generasjon, liksom den gjorde med den andre. Igjen ble feil introdusert i 'kopieringen' og strengen som matchet målsekvensen best, ble valgt som grunnlag for 4.generasjon. Han ga flg. eks. på sine simuleringer: Den tilfeldige startstrengen på 28 tegn var: WDLTMNLT DTJBKWIRZREZLMQCO P. Etter 10 simuleringer fikk han: MDLDMNLS ITJISWHRZRES MECS P. Etter 20 generasjoner: MELDINLS IT ISWPRKE Z WECSEL. Etter 30 generasjoner: METHINGS IT ISWLIKE B WECSEL. Etter 40 generasjoner: METHINKS IT IS LIKE I WEASEL. Og etter 43 generasjoner fikk han målsekvensen: ME THINKS IT LOOKS LIKE A WEASEL. Dawkins kjørte simuleringen flere ganger og fant at han trengte fra 40 til 65 generasjoner for å komme til målstrengen. Dawkins erkjente at modellen hans ikke nøyaktig gjenga hvordan levende organismer fungerte. Han erkjente også at 'til en viss grad er modellen han brukte villedende på viktige områder'. Den ene villedende mangelen som han erkjente, var at seleksjonskriteriet var basert på et langsiktig mål: 'Å komme til målsekvensen'. Kriteriet for naturlig seleksjon, derimot, må være kortsiktig og umiddelbart. Strengen som selekteres på hvert stadium må i seg selv være 'adaptiv'.
 Vi har allerede bemerket at en faktor som er ansvarlig for den sakte farten til evolusjon, er den lave raten til DNA-mutasjoner. Det må også sies at gode mutasjoner er sjeldne, begrepet 'konvergent evolusjon' er behandlet tidligere. Videre har vi vært inne på at selv om en god mutasjon skulle inntreffe, er det stor sjanse for at den vil forsvinne ut fra tilfeldige effekter. Disse faktorene til sammen bidrar til å hindre NDT å fungere i praksis. Dawkins simuleringsmodell i 'The Blind Watchmaker' er selvsagt kunstig, og er ikke bundet av krav til levende organismer. Mutasjonsraten i hans simulering kunne være mye høyere enn hva som kan eksistere i levende liv, uten at populasjoner går til grunne. I levende liv er genomet mye større enn hva Dawkins benyttet i sin simulering. Om mutasjonsraten ikke er liten, så kan en miste det som allerede er bygd opp. Mens Dawkins benytter en mutasjonsrate på 1/28, så er det i naturen snakk om en mutasjonsrate på 1: 10 milliarder. Dawkins valg av høy mutasjonsrate er en av grunnene til at simuleringen fungerer, men den representerer ikke livet. Vi kan dermed ikke forvente at simuleringen skal gi oss noen innsikt i hvordan evolusjon fungerer.
Vi har allerede bemerket at en faktor som er ansvarlig for den sakte farten til evolusjon, er den lave raten til DNA-mutasjoner. Det må også sies at gode mutasjoner er sjeldne, begrepet 'konvergent evolusjon' er behandlet tidligere. Videre har vi vært inne på at selv om en god mutasjon skulle inntreffe, er det stor sjanse for at den vil forsvinne ut fra tilfeldige effekter. Disse faktorene til sammen bidrar til å hindre NDT å fungere i praksis. Dawkins simuleringsmodell i 'The Blind Watchmaker' er selvsagt kunstig, og er ikke bundet av krav til levende organismer. Mutasjonsraten i hans simulering kunne være mye høyere enn hva som kan eksistere i levende liv, uten at populasjoner går til grunne. I levende liv er genomet mye større enn hva Dawkins benyttet i sin simulering. Om mutasjonsraten ikke er liten, så kan en miste det som allerede er bygd opp. Mens Dawkins benytter en mutasjonsrate på 1/28, så er det i naturen snakk om en mutasjonsrate på 1: 10 milliarder. Dawkins valg av høy mutasjonsrate er en av grunnene til at simuleringen fungerer, men den representerer ikke livet. Vi kan dermed ikke forvente at simuleringen skal gi oss noen innsikt i hvordan evolusjon fungerer.
Bilde 5. Mutasjoner og naturlig seleksjon må samvirke
I Dawkins simulerte verden kunne han sørge for at forutsetningene som NDT er avhengig av, holdt:
1. Forutsetningen om forekomst av mutasjoner med positiv seleksjonsverdi sikres ved å ha et 'positivt mål' å sammenligne med. Dermed er en sikret at dette kravet holder, selv om målsekvensen i simuleringen ikke har noe med virkeligheten å gjøre.
2. Mutasjonsraten Dawkins benyttet var høy, og seleksjonen var umiddelbar og sikker -ut fra nevnte målsekvens. Da var ikke forutsetning ii) i NDT nødvendig for å få til en positiv utvikling. I tidligere eks. med evolusjon av hester, var mutasjonsraten som nevnt 1: 10 milliarder pr. nukleotide pr. replikasjon. Videre var det sannsynlig at tilfeldige variasjoner ville fjerne evt. gode mutasjoner, om ikke potensialet for tilpassede mutasjoner var 1 million adaptiv mutasjoner på hvert steg, for at i det minste én av dem skulle inntreffe og overleve. I simuleringen var det en høy frekvens av positive mutasjoner, og Dawkins var sikker på å få minst én positiv mutasjon pr. steg. Videre selekterte han slik at det sikret overlevelse av gode mutasjoner (i forhold til målsekvensen). Det ble ikke overlatt til tilfeldigheter. Det kan beregnes at simulering under slike forutsetninger vil fungere.
Om han hadde kjørt en mer realistisk simulering, ville ha måttet befinne seg lenge ved computeren. Simuleringen ville vært mer realistisk om han hadde benyttet et genom på 500 symboler, i stedet for 28 og en mutasjonsrate på 1: 10 milliarder i stedet for 0,04. Om han hadde benyttet disse realistiske forutsetningene, ville det vært nødvendig med 70 milliarder replikasjoner, bare for å få mutasjonene han trengte. Det ville vært nødvendig at seleksjon til å spre 500 mutasjoner ut gjennom populasjonen. Uten å simulere seleksjon, ignorerer man muligheten for at en god mutasjon vil bli fjernet før den overtar populasjonen.
Bilde 6. Intensitet avtar med antall variable
Dawkins 2. simulering besto av utvikling av linje-tegninger. Han skrev et program for å tegne symmetriske figurer på 'tre-lignende vis', der nye linjer kunne forgrene seg ut fra gamle linjer rekursivt. Han benyttet 9 ulike karakteristika for å definere figurene. Programmet var bygd utfra en symmetri, der han benyttet numeriske verdier fra -9 til +9 for å representere verdiene til de ulike karakteristika. De inkluderte vinkel mellom grenene, lengden av linjestykkene og dybden i rekursjonen.. Han kalte figurene biomorfe, noe som satte ham i stand til ordspill, som om de besto av en populasjon av levende organismer som utviklet seg.
De 9 ulike karakteristika (egenskapene) kunne hver ha 19 ulike verdier. Siden hvert sett av ni karakteristika-verdier representerer en figur, så er det (19 opphøyd i 9) ulike mulige figurer. Det tilsvarer ca. 300 milliarder. Han kunne starte med hvilken som helst figur, og kunne ende opp med hvilken som helst annen, med sitt 'evolusjons-program'. Han sammenliknet karakteristika-verdiene med gener, siden de er arvelige og bestemmer figuren fullstendig. Endring i en karakteristika-verdi spiller mutasjonsrollen i et gen. Simuleringene kopierer en figur, og legger til den minst mulige mutasjon. Det tilsvarer en enhets endring i en av karakteristikaene (enten økning eller reduksjon med 1 enhet). Når verdiene er på topp eller bunn-nivå, så må de hhv. synke eller øke, slik at det blir 18 ulike minste mutasjoner, og dermed 18 ulike figurer som kan bli laget i hvert steg.
(19 opphøyd i 9) ulike mulige figurer. Det tilsvarer ca. 300 milliarder. Han kunne starte med hvilken som helst figur, og kunne ende opp med hvilken som helst annen, med sitt 'evolusjons-program'. Han sammenliknet karakteristika-verdiene med gener, siden de er arvelige og bestemmer figuren fullstendig. Endring i en karakteristika-verdi spiller mutasjonsrollen i et gen. Simuleringene kopierer en figur, og legger til den minst mulige mutasjon. Det tilsvarer en enhets endring i en av karakteristikaene (enten økning eller reduksjon med 1 enhet). Når verdiene er på topp eller bunn-nivå, så må de hhv. synke eller øke, slik at det blir 18 ulike minste mutasjoner, og dermed 18 ulike figurer som kan bli laget i hvert steg.
 Han benyttet en høy mutasjonsrate, som han sa -for: 'mennesker har ikke tålmodighet til å vente en million generasjoner på én mutasjon'. På hvert steg genererte han alle de 18 nærmeste mutantene, og undersøkte utseende på hver av dem. Han gjentok denne prosessen så mange steg som han hadde tålmodighet til å drive med det. Tegningene ble brukt som en følge av vesentlig tre-like grener, men etter diverse kryssinger og re-kryssinger, så lignet de lite på trær. Med innebygd symmetri kan man forestille seg at de har en form, og i noen kan en se noe likt insekter eller fly. Seleksjonen ble utført av operatøren (Dawkins). Simuleringene ble dermed kunstige, mer enn naturlige. Ved å velge figurer som behaget øyet, kunne han få dem til å utvikle seg i ulike retninger. Dawkins tilbød denne simuleringen som et annet eks. på hvordan evolusjon kan fungere.
Han benyttet en høy mutasjonsrate, som han sa -for: 'mennesker har ikke tålmodighet til å vente en million generasjoner på én mutasjon'. På hvert steg genererte han alle de 18 nærmeste mutantene, og undersøkte utseende på hver av dem. Han gjentok denne prosessen så mange steg som han hadde tålmodighet til å drive med det. Tegningene ble brukt som en følge av vesentlig tre-like grener, men etter diverse kryssinger og re-kryssinger, så lignet de lite på trær. Med innebygd symmetri kan man forestille seg at de har en form, og i noen kan en se noe likt insekter eller fly. Seleksjonen ble utført av operatøren (Dawkins). Simuleringene ble dermed kunstige, mer enn naturlige. Ved å velge figurer som behaget øyet, kunne han få dem til å utvikle seg i ulike retninger. Dawkins tilbød denne simuleringen som et annet eks. på hvordan evolusjon kan fungere.
Bilde 7. Skisser tegnet ut fra Dawkins 'evolusjons-program'
Men heller ikke i denne simuleringen, fikk Dawkins tak i poengene som er viktig for å avgjøre hvorvidt evolusjon kan fungere. Simuleringen mangler to viktige trekk:
i) Det er ingen mutasjoner som gjør at evolusjonen 'stopper opp' -ingen dødelige mutasjoner. Bare operatøren beslutter hvorvidt figuren er reproduktiv eller ikke. Det blir dermed ingen begrensninger i mutasjonsraten en kan benytte, i det en tillater at simuleringen er så rask som en ønsker.
ii) Det er veldig mange tilpasningsdyktige mutasjoner. Enhver figur kan være tilpasningsdyktig. Det avhenger bare av operatørens valg og smak.
 I det virkelige liv, er en høy mutasjonsrate skadelig for organismen, og en høy mutasjonsrate kan ødelegge populasjonen. Ett av problemene med evolusjon i det virkelige liv, er at mutasjoner skjer for sjelden, og at det dermed tar veldig lang tid. Når Dawkins speeder opp farten tilstrekkelig, så dekker han over hovedproblemet som gjør den vanskelig for ikke å si umulig. Kumulativ seleksjon fungerer ikke, fordi lav mutasjonsrate gjør sjansen for å få en god mutasjon for liten. Jfr. ovenfor nevnte feil med å tro at jordas alder er lang nok til at hva som helst kan hende. Det er en annen grunn til at evolusjon virket i simuleringen, men ikke i virkeligheten. Når det er en menneskelig operatør, som ønsker videre utvikling, går han gjerne god for en mutasjon, selv om han på noen tidspunkter ikke ser noen virkelig interessante figurer. Men om vi erstattet operatøren med en algoritme, så har simuleringen en sjanse til å feile. Om operatøren hadde fulgt faste regler i seleksjonen, kunne han ha nådd et stadium der ingen av figurene han så ville tilfredsstille kriteriene for seleksjon. Dermed ville evolusjonsprosessen stoppe opp. Men en menneskelig operatør ville ha en tendens til å gjøre reglene fleksible og således kunne holde på videre.
I det virkelige liv, er en høy mutasjonsrate skadelig for organismen, og en høy mutasjonsrate kan ødelegge populasjonen. Ett av problemene med evolusjon i det virkelige liv, er at mutasjoner skjer for sjelden, og at det dermed tar veldig lang tid. Når Dawkins speeder opp farten tilstrekkelig, så dekker han over hovedproblemet som gjør den vanskelig for ikke å si umulig. Kumulativ seleksjon fungerer ikke, fordi lav mutasjonsrate gjør sjansen for å få en god mutasjon for liten. Jfr. ovenfor nevnte feil med å tro at jordas alder er lang nok til at hva som helst kan hende. Det er en annen grunn til at evolusjon virket i simuleringen, men ikke i virkeligheten. Når det er en menneskelig operatør, som ønsker videre utvikling, går han gjerne god for en mutasjon, selv om han på noen tidspunkter ikke ser noen virkelig interessante figurer. Men om vi erstattet operatøren med en algoritme, så har simuleringen en sjanse til å feile. Om operatøren hadde fulgt faste regler i seleksjonen, kunne han ha nådd et stadium der ingen av figurene han så ville tilfredsstille kriteriene for seleksjon. Dermed ville evolusjonsprosessen stoppe opp. Men en menneskelig operatør ville ha en tendens til å gjøre reglene fleksible og således kunne holde på videre.
Foruten det som er nevnt ovenfor, vet ingen om organismer i virkelig liv kan mutere gjennom mange steg, der mutasjonen i hvert steg er endring i et enkelt basepar -og samtidig være tilpasningsdyktig. Videre eksisterer ikke eksempler på mutasjoner som øker informasjon i genomet (se kap. 5). Dawkins simuleringer har dermed ikke direkte relevans for organisk evolusjon, og simulering av linjetegninger kan ikke tjene til å demonstrere evolusjon. Ut fra måten den er bygd opp på, fungerer simuleringen på siden av hvordan evolusjon fungerer -og ikke, i det virkelige liv. Mens vi tidligere har begrunnet hvorfor kumulativ seleksjon ikke fungerer, så gir ikke Dawkins noen begrunnelse for hvorfor  det fungerer. Heller ikke hans håndtering av livets opprinnelse, bygger noen overbevisende sak for et naturlig opphav. Dawkins presenterte bare problemet, uten å løse det. Han beskrev et forslag til livets opprinnelse, men han ga ingen nyttig kvantitativ informasjon i sakens anledning. Og sist for simuleringene, så kastet han barnet ut med badevannet, da han forsøkte å holde simuleringene så enkle at de kunne fungere. Han droppet dermed faktorene som hindrer kumulativ seleksjon fra å fungere i det virkelige liv.
det fungerer. Heller ikke hans håndtering av livets opprinnelse, bygger noen overbevisende sak for et naturlig opphav. Dawkins presenterte bare problemet, uten å løse det. Han beskrev et forslag til livets opprinnelse, men han ga ingen nyttig kvantitativ informasjon i sakens anledning. Og sist for simuleringene, så kastet han barnet ut med badevannet, da han forsøkte å holde simuleringene så enkle at de kunne fungere. Han droppet dermed faktorene som hindrer kumulativ seleksjon fra å fungere i det virkelige liv.
Bilde 8. Operatøren ønske kan påvirke resultatet
Kort sagt er NDT en dårlig basis for Dawkins tro på et naturlig opphav til livet {noe som kan være en grunn til at han senere har modifisert seg noe her-oversetters kommentar}. Dette var hva han anga som grunn for ikke å tro på noen skaper og herre over universet. Det er synd at en slik dyptgående konklusjon skal baseres på en dårlig understøttet teori (NDT). Det er fare for at bekreftelsesavvik (confirmation bias) gjorde at han fulgte følelser i stedet for fornuft.. I følge papiromslaget konkluderer Dawkins med at "Det kan være gode grunner for tro på en G-d, men argumentet fra design (AFD) er ikke ett av dem". Forfatteren her konkluderer annerledes: "Det kan være .. grunner for å være en ateist, men NDT om evolusjon er ikke én av dem".
Bilde 9. NDT gir ikke grunnlag for livets opprinnelse
{Sluttord: Det er etter hvert en kjent sak at Richard Dawkins hadde et lettere slagtilfelle i starten av febr. 2016. Til tross for Dawkins iherdige ateistiske propaganda, har det blitt for mye' for enkelte. Det har vært rapportert tilfeller av at folk er blitt kristne grunnet hans overdrivelser, og at han er kommet på bedre tanker ang. kristendom. Vi får ønske ham god bedring, uten å være så 'insensitive' at vi kunngjør at vi vil be for ham -oversetters kommentar.}
Nevnte kilder:
1. 'Born in a watery commune; Nature, 427, s.674-676
2: 'Born in a watery commune; Nature, 427, s.26ff.
For øvrig kildematerial henvises til 'Not By Chance; kap. 6, av Dr. Lee Spetner.
Oversettelse samt utvalg av stoff og bilder ved Asbjørn E. Lund